ВЗАЄМОДІЯ НЕАЛЕЛЬНИХ ГЕНІВ




Під взаємодією неалельних генів у вузькому значенні зазвичай розуміють випадки, коли декілька різних генів впливають на розвиток однієї
ознаки. При цьому також спостерігаються відхилення від очікуваних
співвідношень фенотипових класів. Слід розуміти, що йдеться не про
безпосередню взаємодію між генами: прояв ознаки залежить від продуктів
різних генів (наприклад, ферментів, які залучені до одного біохімічного
каскаду). Розрізняють три типи взаємодії неалельних генів: комплементарність, епістаз і полімерію.
Комплементарність
- тип взаємодії неалельних генів, при якому присутність у зиготі
домінантних алелів цих генів (хоча б по одному для кожного гена)
зумовлює такий прояв ознаки, який не спостерігається за наявності лише
одного з домінантних алелів окремо. Гени, що взаємодіють за такою
схемою, називають комплементарними.
Конкретний вид розщеплень на фенотипові класи за комплементарної
взаємодії залежить від того, чи мають домінантні алелі кожного з генів
власний фенотиповий прояв.
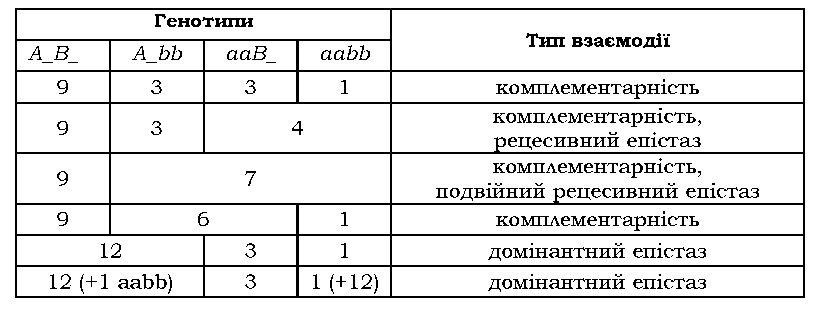
Розглянемо найпростіший випадок взаємодії двох комплементарних генів A/ a і В/ b. Згідно з третім законом Менделя, при дигібрид-ному схрещуванні очікуваним розщепленням за фенотипом у F2 є 9А_В_ : 3А_ЬЬ : 3aaB_ : 1aabb.
Саме це розщеплення і спостерігається при спадкуванні форми гребня в
курей (рис. 3.4), коли кожен із домінантних алелів двох комплементарних
генів має свій власний фенотиповий прояв (табл. 3.3).
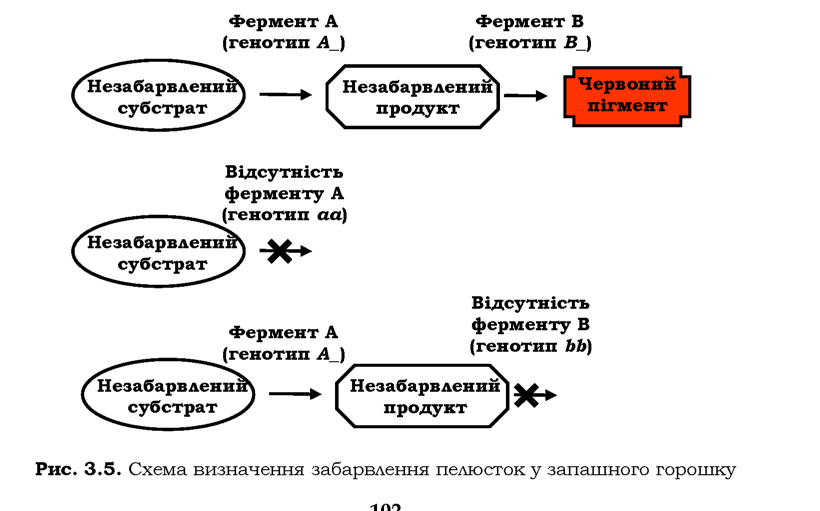
У
запашного горошку забарвлення квітки в червоний колір залежить від двох
домінантних алелів, жоден із яких не має свого власного фенотипового
прояву. Гени кодують два ферменти, залучені до послідовного
двоступеневого перетворення певного субстрату на червоний пігмент: у
відсутності хоча б одного з ферментів (хоча б одного домінантного алеля)
пелюстки квітки залишаються незабарвленими (рис. З.5). Тобто генотипові
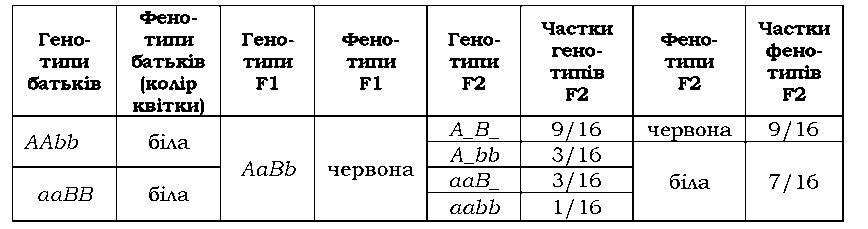
класи А_ЬЬ, aaB_ і aabb мають однаковий фенотип (білі пелюстки) і формують один фенотиповий клас: у F2 спостерігається розщеплення 9 : 7 (табл. З.4).
Таблиця 3.4. Комплементарна взаємодія генів, які визначають забарвлення квітки в запашного горошку
|
 |
Забарвлення
плоду в гарбуза також визначається двома генами (за схемою, аналогічною
до такої на рис. 3.5), але тільки один із домінантних алелів цих генів
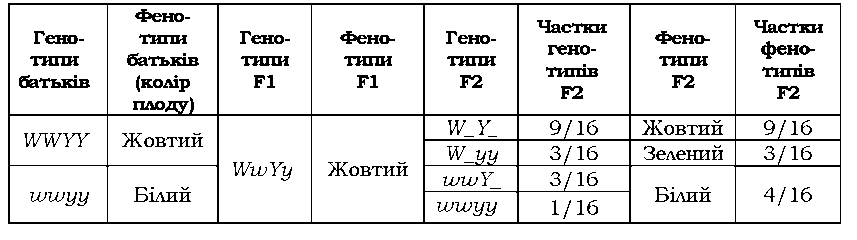
не має власного прояву - у F2 спостерігається розщеплення 9 : 3 : 4
(табл. 3.5). У даному випадку алель W - один із генів, який відповідає за синтез або накопичення зеленого пігменту (хлорофілу). У присутності алеля Y зелений пігмент перетворюється на жовтий (тобто, алель Y у фенотипі виявляється виключно за наявності W, але власного фенотипового прояву не має). У присутності двох алелів w пігмент узагалі не утворюється, тому плоди залишаються білими.
Таблиця 3.5. Комплементарна взаємодія генів, які визначають забарвлення плоду в гарбуза
|
 |
Ще
один тип розщеплення у другому поколінні при комплементарній взаємодій
генів спостерігається тоді, коли обидва домінантні алелі мають однаковий
власний фенотиповий прояв. Так, домінантні алелі двох певних генів
визначають однакову сферичну форму плоду в гарбуза, якщо вони присутні в
генотипі окремо; сумісна дія цих алелів зумовлює дископодібну форму. У
другому поколінні спостерігаються три фенотипові класи у співвідношенні 9
: 6 : 1 за рахунок об'єднання генотипових класів A_bb і aaB_ (табл. 3.6).
Епістаз - тип взаємодії неалельних генів, при якому один ген здатен пригнічувати дію іншого. Ген, який пригнічує, називають епі-статичним, а ген, дія якого пригнічується, - гіпостатичним.
Характер розподілу фенотипових класів залежить від наявності власного
прояву у двох генів, що взаємодіють, та від того, домінантний чи
рецесивний алель гена є епістатичним.
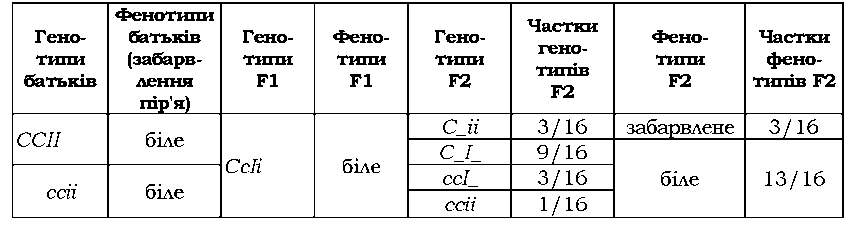
Якщо інгібітором є домінантний алель епістатичного гена, має місце домінантний епістаз (табл. З.7 - алель І пригнічує прояв алеля С при забарвленні пір'я в курей).
Таблиця 3.7. Епістатична взаємодія генів, які визначають забарвлення пір'я в курей
|
 |
Іншим
варіантом розщеплення при домінантному епістазі може бути розщеплення
12 : З : 1. Це спостерігається при забарвленні хутра в деяких гризунів.
Алель А зумовлює синтез чорного пігменту, а алель В
-рудого. За відсутності обох домінантних алелів хутро залишається
білим. За наявності двох алелів (А_В_) синтезуються обидва пігменти, але
рудий маскується чорним - хутро в цьому разі має чорне забарвлення.
У
випадку, коли гіпостатичний ген не виявляється за присутності
епістатичного рецесивного алеля в гомозиготному стані, говорять про рецесивний епістаз.
Як приклад даного типу взаємодії можна навести спадкування кольору
шерсті в лабрадорських мисливських собак. Один ген зумовлює тип
пігменту, який синтезується клітинами шкіри: алель B зумовлює синтез чорного пігменту, а рецесивний алель b -коричневого. Інший ген контролює накопичення пігменту у волоссі: наявність домінантного алеля E приводить до накопичення пігменту, тоді як у гомозигот за рецесивниим алелєм e пігменту у волоссі немає, такі собаки мають жовтий колір. Отже, алель e
в гомозиготному стані приводить до відсутності пігменту у волоссі,
тобто можна сказати, що він пригнічує прояв гена забарвлення. Очевидно,
що генотипові класи B_ee та bbee будуть об'єднуватися в один фенотиповий, і при схрещуванні дигетерозигот спостерігатиметься розщеплення 9 (B_E_ - чорні) : 3 (bbE_- коричневі) : 4 (3 B_ee + 1 bbee
- жовті). Очевидно також, що наведений приклад рецесивного епістазу
скоріше можна інтерпретувати як комплементарність: чорне забарвлення
волосся спостерігається тільки за наявності домінантних алелів обох
генів (див. аналогічне розщеплення в табл. 3.5).
Поряд із простим рецесивним епістазом, іноді формально розглядають подвійний рецесивний епістаз
- ситуацію, коли рецесивний алель будь-якого з генів, що взаємодіють, у
гомозиготному стані є епі-статичним щодо іншого гена. Так, розглянуте
розщеплення 9 : 7 (спадкування забарвлення квітки в запашного горошку,
табл. 3.4) можна інтерпретувати як подвійний рецесивний епістаз: aa
пригнічує прояв алеля B, а bb, у свою чергу, - прояв A. Наведені
приклади демонструють, що рецесивний епістаз (простий або подвійний)
насправді зводиться до комплементарності.
Чітко
визначити тип взаємодії неалельних генів можливо в тих випадках, коли
відомий біохімічний механізм прояву досліджуваної ознаки (як, скажімо, у
випадку забарвлення квітки в запашного горошку). Часто біохімічний
механізм залишається невідомим, і тоді обмежуються констатацією
формальної генетичної схеми спадкування ознаки. Отже, ідентифікація того
чи іншого типу взаємодії неалельних генів часто є умовною. Як показано в
табл. 3.3-3.7, при відхиленнях у розщепленнях, зумовлених взаємодією
двох генів, співвідношення фенотипових класів, що спостерігаються у F2,
завжди можна вивести з класичного 9 : 3 : 3 : 1. Для цього важливо
усвідомити, які саме класи об'єднуються, після чого можна зробити
висновок про можливий тип взаємодії (табл. 3.8).
Таблиця 3.8. Співвідношення фенотипових класів у F2 дигібридного схрещування при деяких типах взаємодії неалельних генів
|
 |
Полімерія - тип взаємодії неалельних генів, при якому ознака формується в результаті дії кількох генів з однаковим впливом на ознаку (однозначних генів),
тобто йдеться про родини ідентичних або гомологічних генів, що
повторюються в геномі та мають однаковий прояв. Сумарний ефект може
залежати просто від наявності хоча б одного такого гена (при визначенні
якісних ознак) або від кількості домінантних алелів взаємодіючих генів
(при визначенні кількісних ознак). Відповідно, розрізняють некумулятивну та кумулятивну
полімерію. У разі полімерії прийнято позначати взаємодіючі гени однією
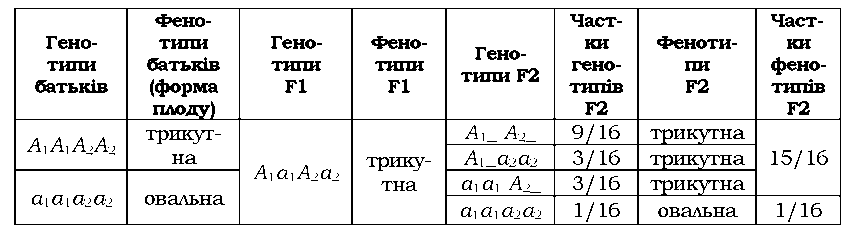
буквою, різні гени при цьому позначають нижнім цифровим індексом (А1а1А2а2Аэа3...). При некумулятивній полімерії
достатньо одного домінантного алеля будь-якого гена з числа тих, які
впливають на ознаку, щоб ця ознака виявилась. Характерне розщеплення у
другому поколінні 15 : 1, прикладом є форма плоду у грициків Capsella bursa-pastoris (табл. 3.9).
Таблиця 3.9. Полімерна взаємодія генів, які визначають форму плоду
в Capsella. bursa-pastoris
|
 |
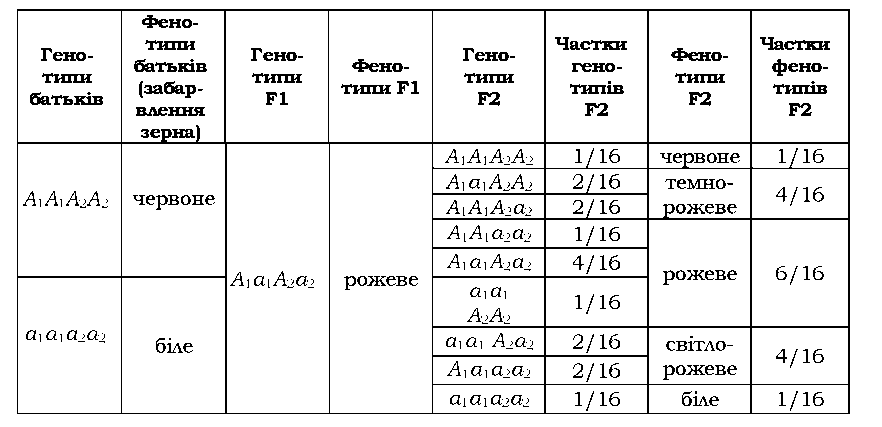
У випадку кумулятивної полімерії
ступінь прояву ознаки (яку називають адитивною) залежить від кількості
домінантних алелів полімерних (адитивних) генів: можна сказати, що вплив
окремих генів накопичується, і чим більше домінантних алелів різних
генів, тим сильніше проявляється ознака. Саме за таким принципом
спадкується, наприклад, колір шкіри в людини (рис. 3.6).
Для
двох пар адитивних генів характерне розщеплення у другому поколінні
становить 1 : 4 : 6 : 4 : 1 - прикладом є розвиток забарвлення зерна
пшениці (табл. 3.10). За типом кумулятивної полімерії спадкується
більшість кількісних ознак.
Таблиця 3.10. Полімерна взаємодія генів, які визначають забарвлення зерна пшениці
|
 |
Немає коментарів:
Дописати коментар